

Электрофизиология
Механорецептивные волосковые клетки структуры боковой линии интегрируются в более сложные цепи через свои афферентные и эфферентные связи. Синапсы, которые непосредственно участвуют в передаче механической информации, представляют собой возбуждающие афферентные связи, использующие глутамат . Однако возможно множество различных нейромастов и афферентных связей, что приводит к изменению механорецептивных свойств. Например, серия экспериментов на поверхностных невромастах Porichthys notatus показала, что невромасты могут проявлять рецептивную специфичность для определенных частот стимуляции. Используя обездвиженную рыбу для предотвращения посторонней стимуляции, металлический шар вибрировал с разной частотой. Используя измерения отдельных клеток с помощью микроэлектрода, ответы были записаны и использованы для построения кривых настройки, которые выявили частотные предпочтения и два основных типа афферентных нервов. Одна разновидность приспособлена для сбора механорецептивной информации об ускорении, реагируя на частоты стимуляции от 30 до 200 Гц. Другой тип чувствителен к информации о скорости и наиболее восприимчив к стимуляции ниже <30 Гц. Это предполагает более сложную модель приема, чем считалось ранее.
Эфферентные синапсы к волосковым клеткам тормозят и используют ацетилхолин в качестве передатчика. Они являются важными участниками системы разряда следствий, предназначенной для ограничения самогенерируемых помех. Когда рыба движется, она создает волнения в воде, которые могут быть обнаружены системой боковой линии, потенциально препятствуя обнаружению других биологически значимых сигналов. Чтобы предотвратить это, при двигательном действии волосковой клетке посылается эфферентный сигнал, что приводит к торможению, которое противодействует возбуждению, возникающему в результате приема самогенерируемой стимуляции. Это позволяет рыбе сохранять восприятие стимулов движения без помех, создаваемых ее собственными движениями.
После того, как сигналы, передаваемые волосковыми клетками, передаются по боковым нейронам, они в конечном итоге достигают мозга. Методы визуализации показали, что область, где эти сигналы чаще всего заканчиваются, – это медиальное октаволатеральное ядро (MON). Вероятно, что MON играет важную роль в обработке и интеграции механорецептивной информации. Это было подтверждено другими экспериментами, такими как использование окрашивания по Гольджи и микроскопия New & Coombs, чтобы продемонстрировать присутствие отдельных слоев клеток в MON. Отчетливые слои базилярных и небазилярных клеток гребня были идентифицированы внутри глубокого MON. Проведя сравнение с аналогичными клетками в близкородственной электросенсорной доле боковой линии электрических рыб, можно предположить возможные вычислительные пути МОН. MON, вероятно, участвует в интеграции сложных возбуждающих и тормозных параллельных цепей для интерпретации механорецептивной информации.
Интересные факты
- Нейромасты родственны нервным клеткам, отвечающим у нас за равновесие. В какой-то момент эволюция пошла по другому пути, и обделила нас сейсмосенсорикой. Впрочем, у некоторых сухопутных животных она имеется, пусть и не в столь развитом состоянии, иначе как объяснить тот факт, что собаки и кошки чувствуют приближение землетрясений?
- Большинство рыб способно улавливать колебания стеклянного волоска толщиной в четверть миллиметра на расстоянии до полуметра! Чувствительность боковой линии зависит от вида рыбы, но абсолютно «инфраглухих» среди них не наблюдается.
- Жерех научился использовать сейсмосенсорику рыб против них же самих. Этот хищник из семейства карповых выработал абсолютно уникальную тактику охоты. Он охотится максимально шумно, выпрыгивая из воды и создавая плеск внушительным хвостом. В результате рыбки-жертвы буквально «глохнут» и дезориентируются в пространстве, а жерех спокойно пожирает растерявшуюся добычу.
- Кефаль и сельдь не имеют боковой линии. То есть, не имеют ее визуального проявления вдоль тела. У этих рыб большинство нейромастов расположено на голове, а также имеются свободные сенсоры на коже.
- Морской окунь-терпуг имеет несколько боковых линий. Природа «украсила» представителей семейства терпуговых несколькими боковыми линиями. И это закономерно: большинство их обитает на значительных глубинах, где свет в дефиците.
Если знать об уникальной сейсмочувствительности рыб, можно избежать множества ошибок и понять, как себя вести на рыбалке в том или ином случае. Богатых уловов вам, коллеги!
Рыболовы удивляются, почему у меня клюет, а у них нет? Только для вас раскрываю секрет: все дело в чудо-приманке! Подробнее
Анатомия
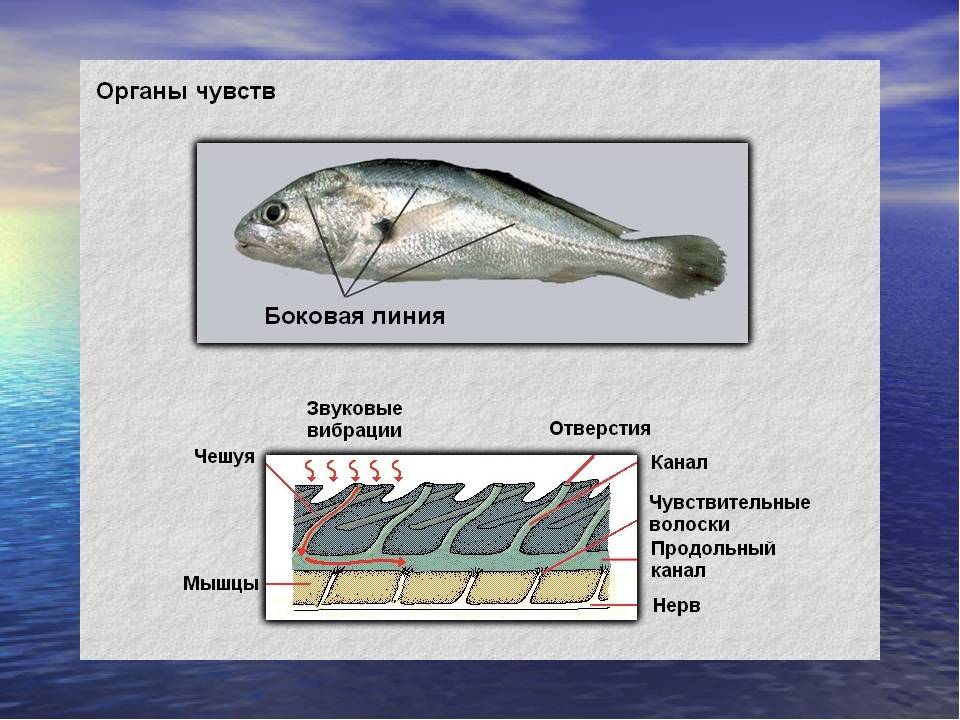
Схема системы боковой линии
Некоторые чешуйки боковой линии (центра) Рутилус рутилус.
А трехиглая колюшка с окрашенными невромастами
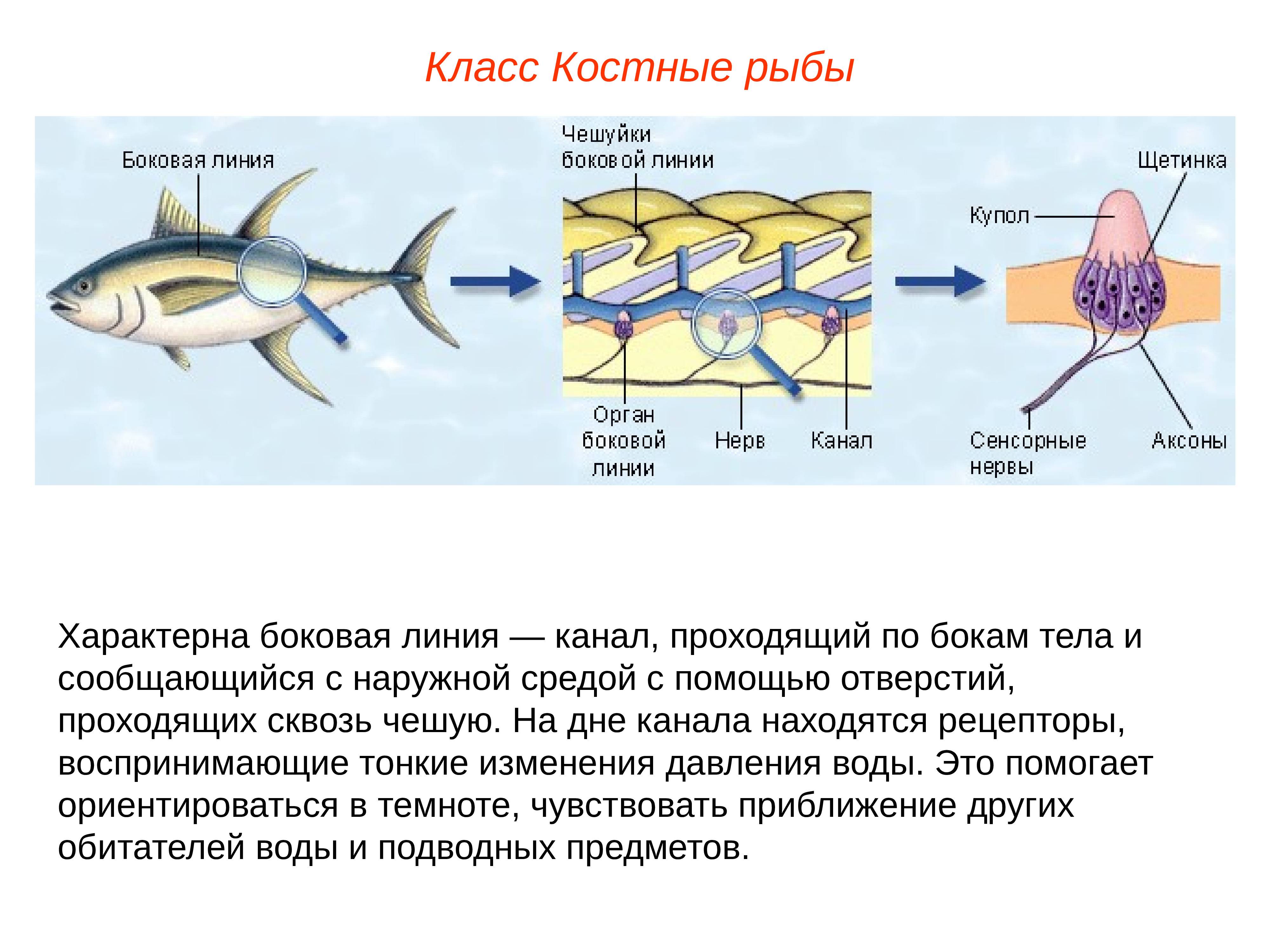
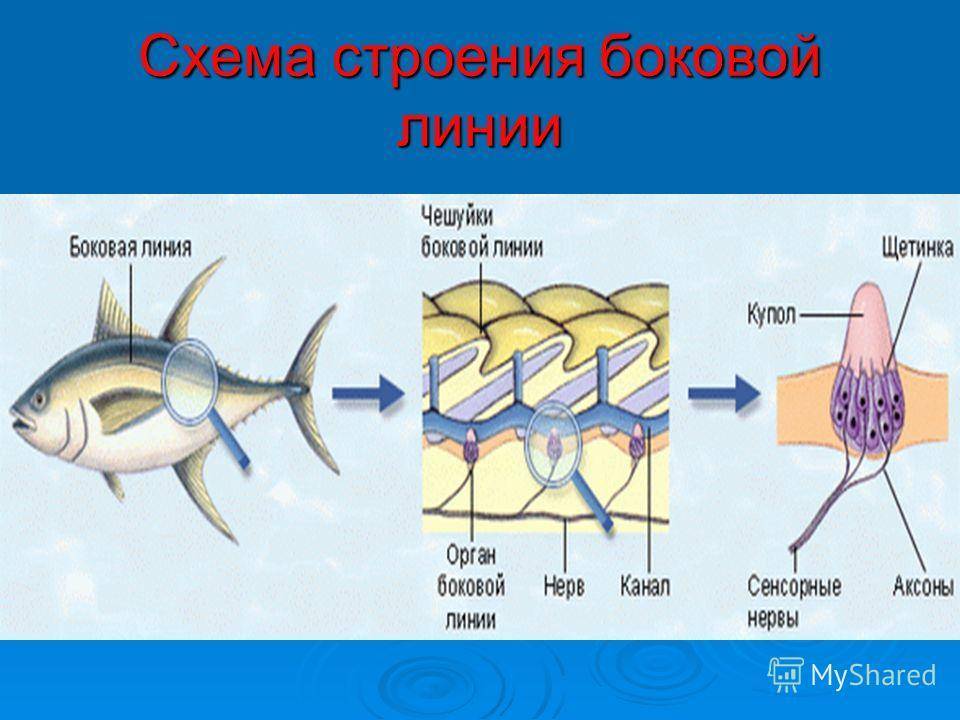
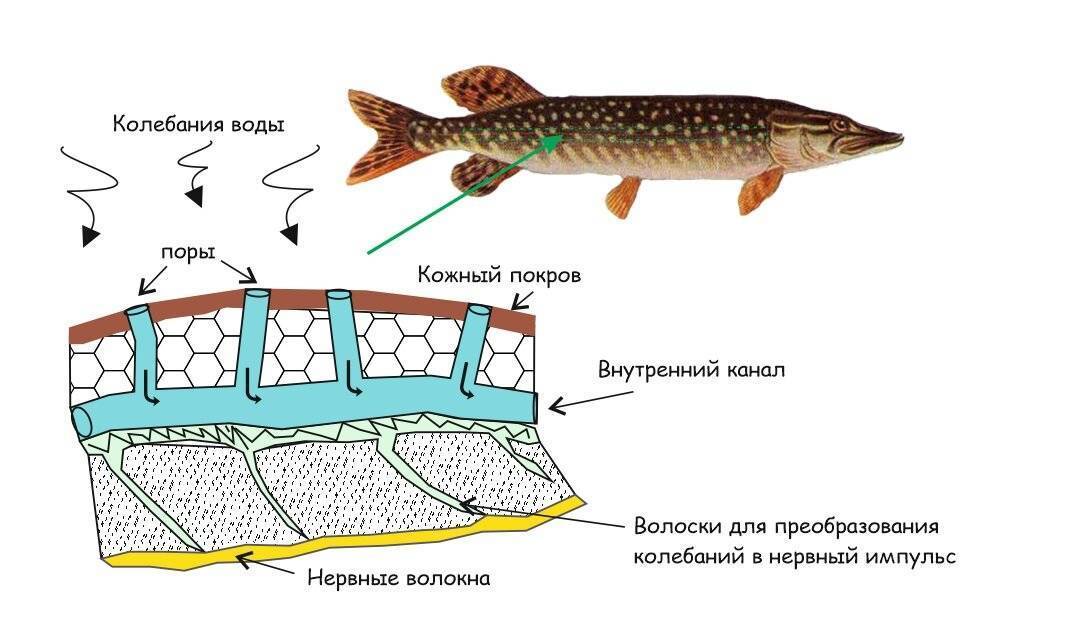
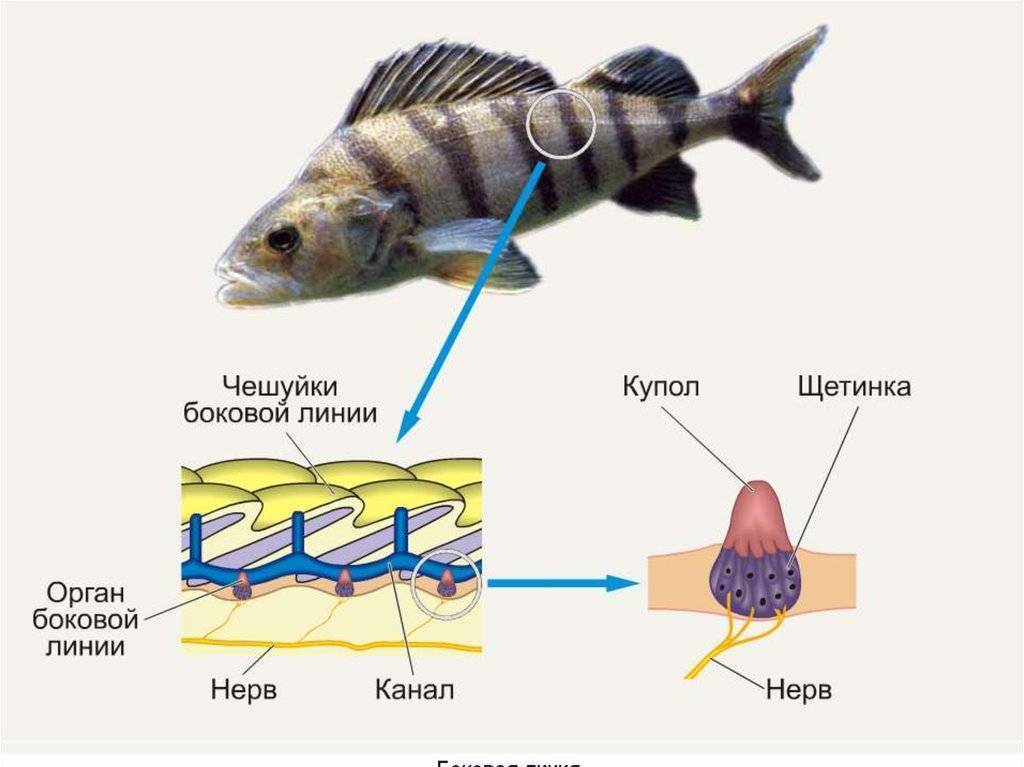
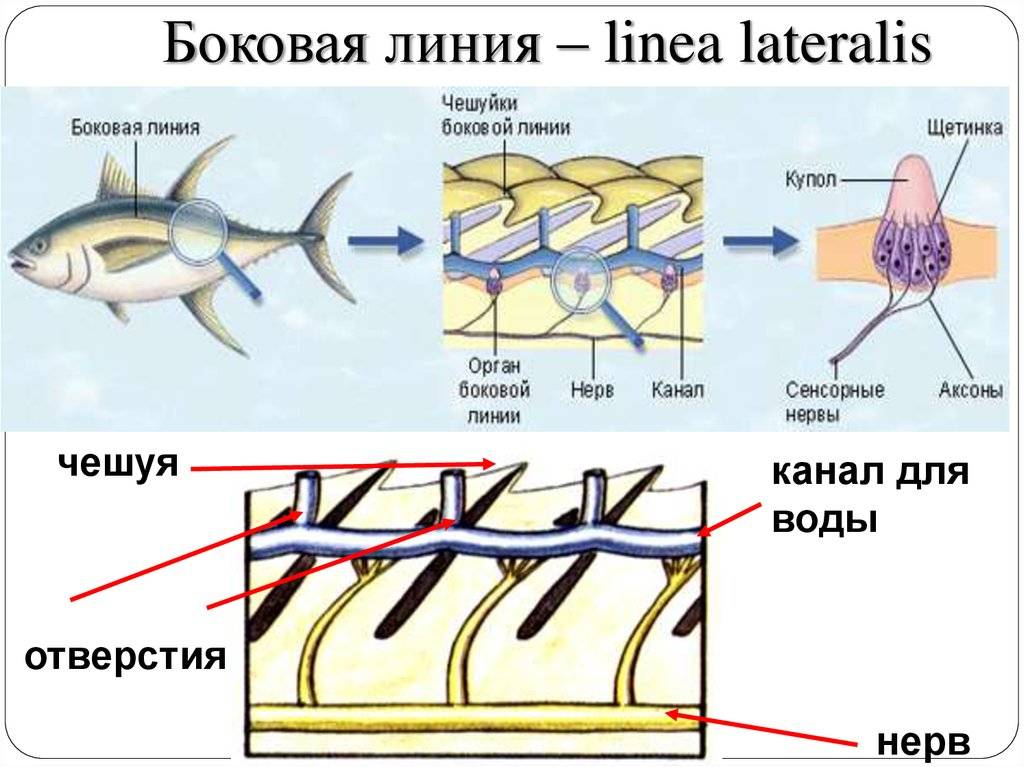
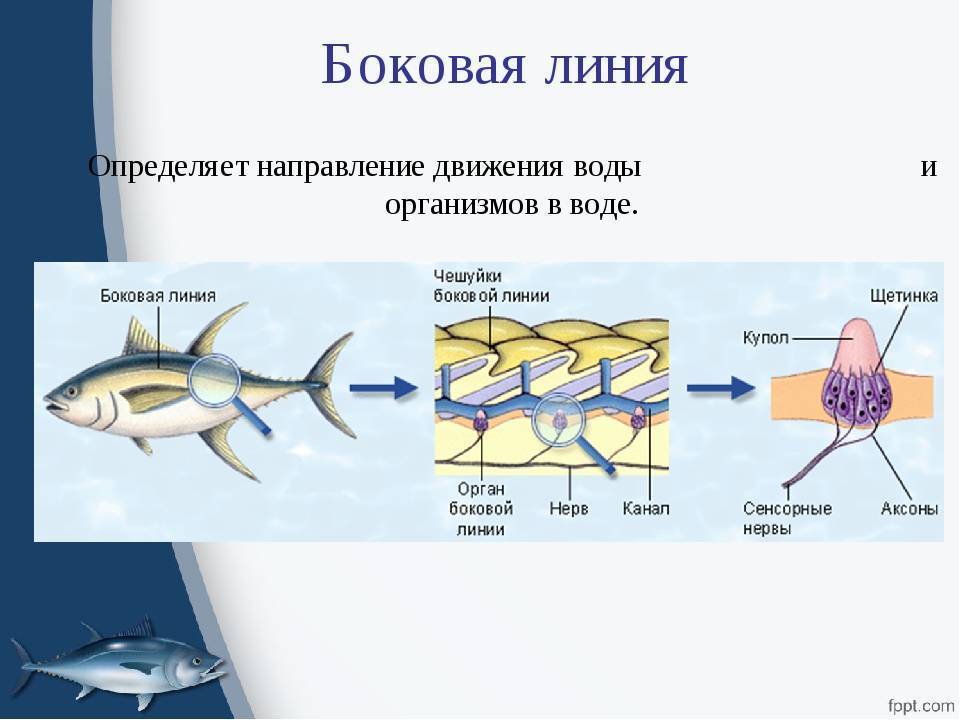
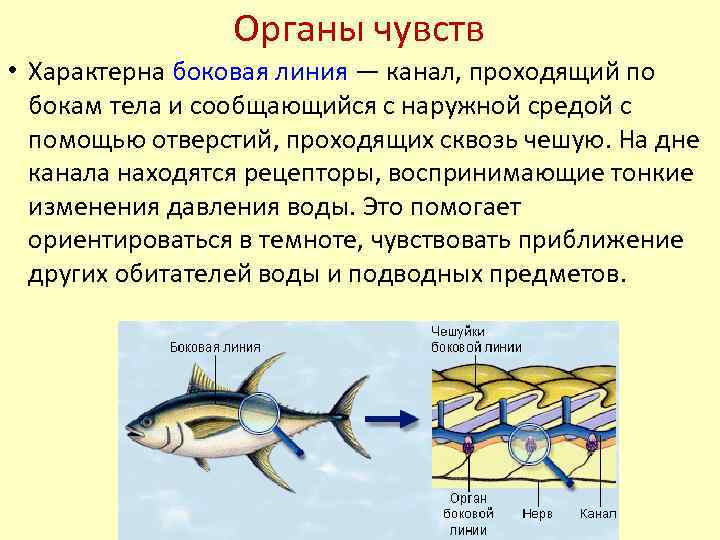
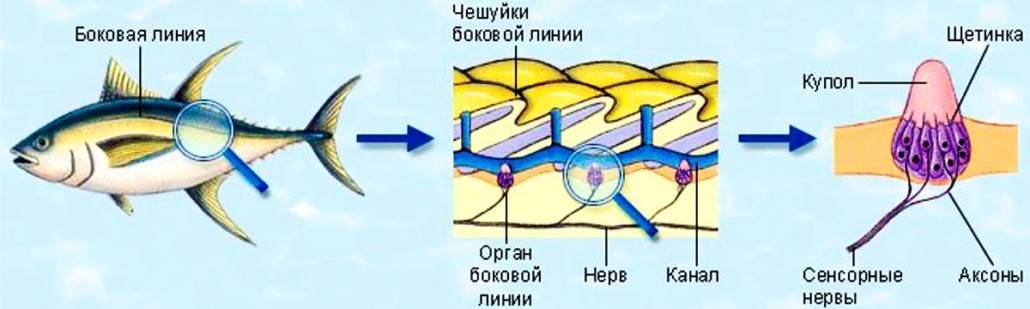
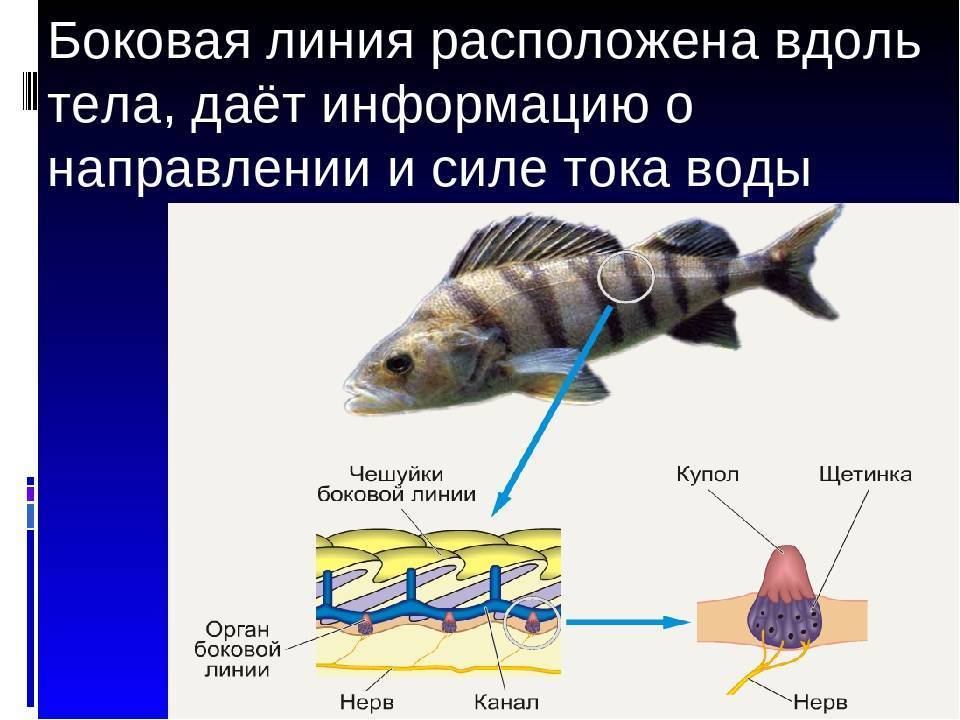
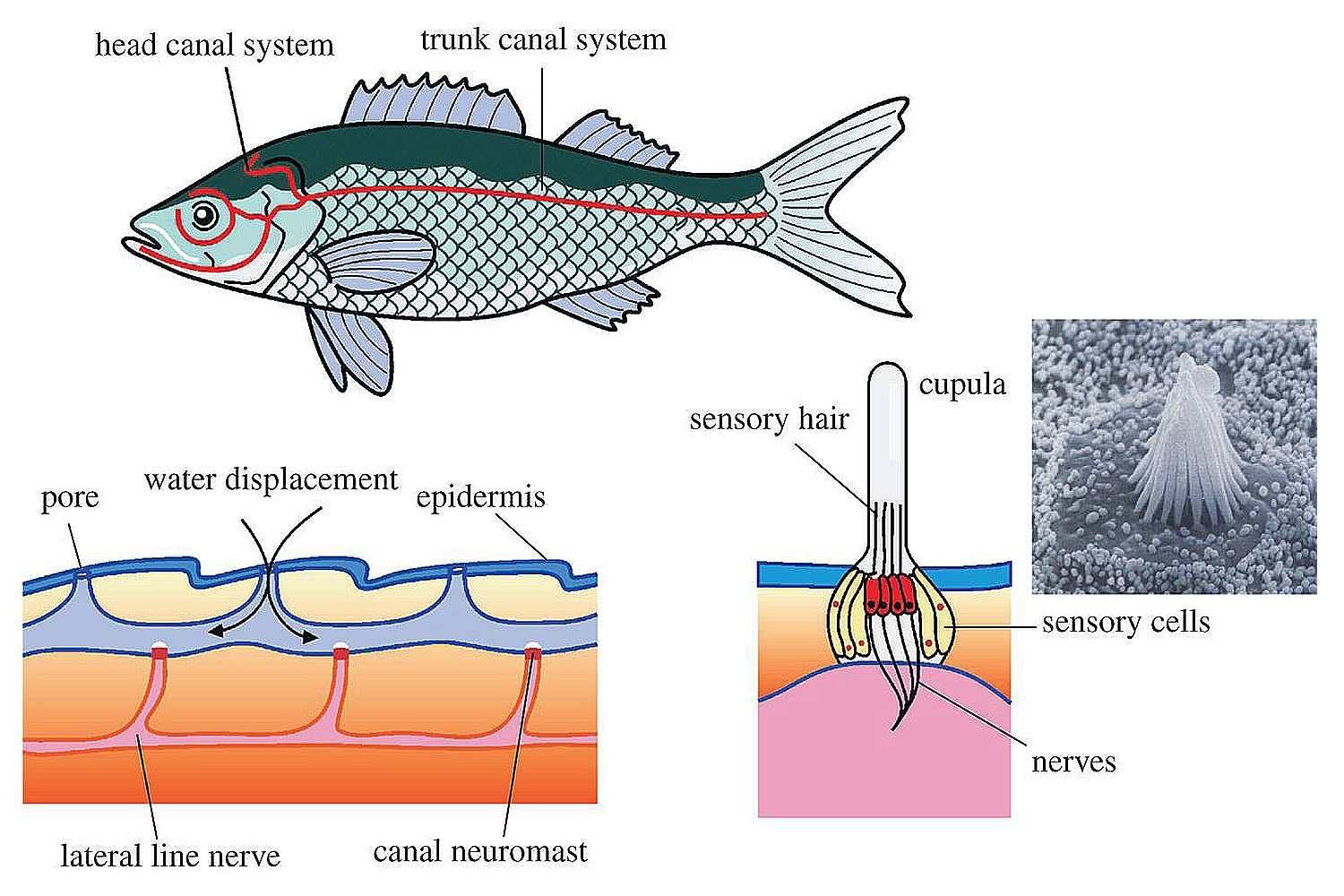
Важнейшей функциональной единицей боковой линии является невромаст. Невромаст – это механорецептивный орган, который позволяет ощущать механические изменения в воде. Существует две основных разновидности невромастов, расположенных у животных: канальные невромасты и поверхностные или отдельно стоящие невромасты. Поверхностные невромасты расположены снаружи на поверхности тела, в то время как невромасты каналов расположены вдоль боковых линий в подкожных каналах, заполненных жидкостью. Каждый невромаст состоит из рецептивных волосковые клетки кончики которых покрыты гибкой студенистой купулой. Волосковые клетки обычно обладают обоими глутаматергический афферентные связи и холинергический эфферентные связи. Изменяются рецептивные волосковые клетки. эпителиальные клетки и обычно имеют связки по 40-50 микроворсинки “волосы”, которые функционируют как механорецепторы. Эти пучки организованы в грубые «лестницы» из волосков увеличивающегося порядка длины. Такое использование механочувствительных волос аналогично функционированию волосковых клеток в слуховой и вестибулярная система, что указывает на тесную связь между этими системами.
Клетки волос использовать систему трансдукция который использует кодирование скорости чтобы передать направленность стимула. Волосковые клетки системы боковой линии производят постоянную тоническую скорость возбуждения. Поскольку механическое движение передается через воду к невромасту, купула изгибается и смещается. Различаясь по величине в зависимости от силы стимула, происходит срезание и отклонение волос либо в сторону самых длинных волос, либо от них. Это приводит к сдвигу ионной проницаемости клетки в результате изменения ее открытия. ионные каналы вызвано отклонением волосков. Отклонение к самым длинным волосам приводит к деполяризация волосковой клетки, повышенное высвобождение нейротрансмиттера в возбуждающем афферентном синапсе и более высокая скорость преобразование сигнала. Отклонение к более коротким волосам имеет противоположный эффект, гиперполяризующий волосковая клетка и снижает скорость высвобождения нейромедиатора. Эти электрические импульсы затем передаются по афферентным латеральным нейронам в мозг.
Хотя обе разновидности невромастов используют этот метод трансдукция, специализированная организация поверхностных и канальных невромастов позволяет им различать механорецептивный мощности. Расположенные на поверхности кожи животного, поверхностные органы более подвержены воздействию внешней среды. Хотя эти органы обладают стандартными пучками волос в форме «лестницы», в целом организация пучков внутри органов кажется случайной, включая различные формы и размеры. микроворсинки в пачках. Это предполагает широкий диапазон обнаружения, потенциально указывающий на функцию широкого обнаружения для определения наличия и величины отклонения, вызванного движением в окружающей воде. Напротив, структура органов каналов позволяет организовывать невромасты каналов в сетевую систему, которая обеспечивает более сложные механорецепция, например, обнаружение перепада давления. Когда ток проходит через поры канала, над порами создается перепад давления. Когда давление на одну пору превышает давление другой поры, перепад давит на канал и вызывает поток жидкости в канале. Это перемещает купулу волосковых клеток в канале, что приводит к направленному отклонению волос, соответствующему направлению потока. Этот метод позволяет преобразовывать информацию о давлении в направленные отклонения, которые могут быть получены и преобразованный волосковыми клетками.
Рост и развитие рыб
Из оплодотворенной икринки развивается личинка, отличающаяся от взрослых рыб формой тела и нередко наличием различных временных органов.
В зависимости от количества желтка желточный мешок сохраняется у личинок более или менее длительный срок. По окончании его рассасывания личинки большинства рыб приобретают внешний вид, характерный для взрослых особей данного вида; они переходят на активное питание и превращаются в мальков. У некоторых видов рыб этот процесс несколько затягивается.
Уже на стадии личинки начинается линейный и весовой рост рыбы. В этот период рост происходит в основном за счет рассасывания желточного мешка. При переходе на активное питание рост продолжается за счет пищи. Рыбы растут в течение всей жизни; вначале этот процесс протекает быстро, затем рост все более замедляется.
Помимо возраста рыбы скорость ее роста зависит от цело-то ряда факторов. Большое значение имеет количество и качество пищи; большинство рыб на более ранних стадиях развития нуждаются в большом количестве сравнительно однообразной пищи, для многих же взрослых рыб желательна смена кормов. Важную роль играет температура; как правило, чем выше температура, тем быстрее рост, доходящий до максимума в оптимальных для данного вида условиях и замедляющийся при дальнейшем повышении температуры, а затем полностью прекращающийся. Гидрохимические условия также влияют на скорость роста рыб. Не последнее место занимает и видовая принадлежность рыбы (рыбы разных видов растут при оптимальных условиях с разной скоростью).
Важно сочетание всех благоприятных факторов. Так, оптимальные температурные и кислородные условия без необходимого количества пищи или указанных факторов при неблагоприятных условиях рН будут недостаточны для нормального роста
При различном сочетании факторов, влияющих на рост может быть несоответствие между увеличением линейного роста (длины) и весом рыбы.
Развитие начинается с момента оплодотворения икринки и делится на два периода — эмбриональный и постэмбриональный; последний начинается после выхода личинки из яйцевых оболочек. Для нормального развития играют роль в основном те же факторы, что и для роста.



Однако значимость отдельных факторов для роста и развития рыб в отдельных случаях не совпадает. Так, при определенном кормлении может происходить нормальный рост, но развитие половой системы задерживается или совершенно прекращается (в результате ожирения). И, наоборот, часто наблюдается более быстрое созревание при задерживающемся росте.
При наступлении половой зрелости окончательно развиваются половые железы, выделяемые ими гормоны вызывают развитие половых признаков. По этим признакам и отличают в большинстве случаев самцов от самок. К первичным половым признакам относятся копулятивные органы самцов живородок и цихлид, яйцеклады самок цихлид,
Развитие с возрастом рыб их половых желез вызывает часто появление вторичных половых различий в размерах тела, величине и форме плавников, очень часто в степени яркости и характере окраски. Многие вторичнополовые признаки в гораздо большей степени проявляются во время размножения; некоторые — только в период размножения (жемчужинки у золотых рыбок), это так называемый брачный наряд.
Большинство аквариумных рыб живут не более 3—5 лет, некоторые цихлиды — свыше 10 лет, золотые рыбки — несколько, десятков лет. Некоторые виды икромечущих карпозубых, живущие в естественных условиях всего лишь несколько месяцев, в аквариуме живут несколько лет. Можно считать, что в аквариумах рыбы, как правило, в благоприятных условиях живут дольше, чем в природных. Здесь они лишены конкуренции и получают достаточное количество пищи. У бойцовых рыб замечено, что они живут значительно меньше при частых помещениях их на нерест.
Размножение
У самцов хрящевых рыб (акул и скатов), а также некоторых живородящих лучеперых рыб, развиты модифицированные плавники, выполняющие роль мужского полового органа, репродуктивных придатков, с помощью которых эти рыбы осуществляют внутреннее оплодотворение. У лучеперых рыб эти органы называются гоноподий и андроподий, у хрящевых рыб — класперами.
 Видоизмененный анальный плавник у самца гуппи — гоноподий
Видоизмененный анальный плавник у самца гуппи — гоноподий
Гоноподий можно обнаружить у некоторых самцов из семействчетырехглазковые и пецилиевые. Это анальные плавники, которые в результате мутаций стали функционировать как подвижные половые органы и используются для оплодотворения самок с помощью молок во время спаривания. Третий, четвертый и пятый лучи анального плавника у самца формируют желобок, по которому движутся сперматозоиды рыбы. Когда наступает момент спаривания, гоноподий выпрямляется и указывает прямо на самку. Вскоре половой орган самца, оснащенный похожим на крюк отростком, входит в половые органы самки. Этот отросток необходим самцу, чтобы удержаться рядом с самкой во время оплодотворения. Если самка сохраняет неподвижность во время этого процесса, оплодотворение проходит успешно. Сперма сохраняется в яйцеводе самки. Это позволяет самке оплодотворить себя в любой момент без дополнительной помощи самца. У некоторых видов длина гоноподия может соответствовать половине общей длины тела. Иногда длина плавника такова, что рыба не способна использовать орган, как это бывает у «лирохвостых» пород зеленых меченосцев. Развитие гоноподия возможно и у самок после приема гормональных препаратов. Однако такие рыбы бесполезны для селекции.
Аналогичные органы с похожими характеристиками встречаются и у других рыбы, например, андроподий у Хемирхамфодона или Гудиевых.
Класперы встречаются у самцов хрящевых рыб. Они расположены на задней части брюшных плавников и тоже в результате изменений стали выполнять функции репродуктивных органов — поставлять сперму в клоаку самки во время спаривания. В процессе спаривания акул один из класторов обычно поднимается, чтобы вода могла проникнуть в сифон через специальное отверстие. Затем кластер входит в клоаку, где он раскрывается наподобие зонтика и закрепляется в определенной позиции. Затем в сифон начинают поступать вытесняемые вода и сперма.
Внешнее строение рыбы
Внешнее строение водных жителей напрямую связано со средой обитания, и кардинально отличается от других классов животных.
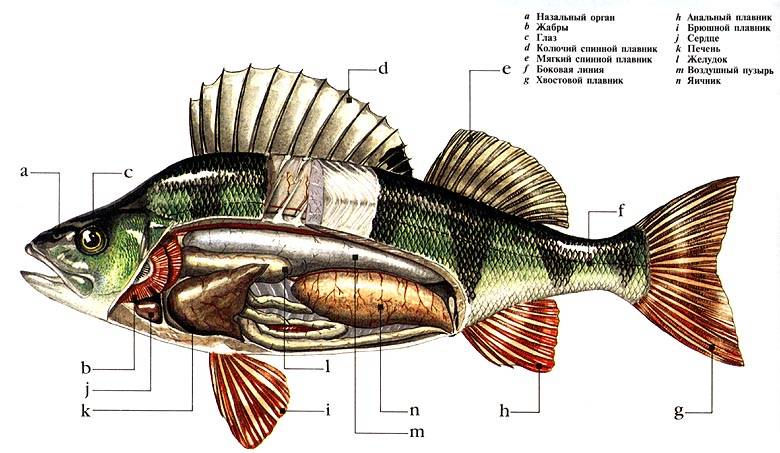
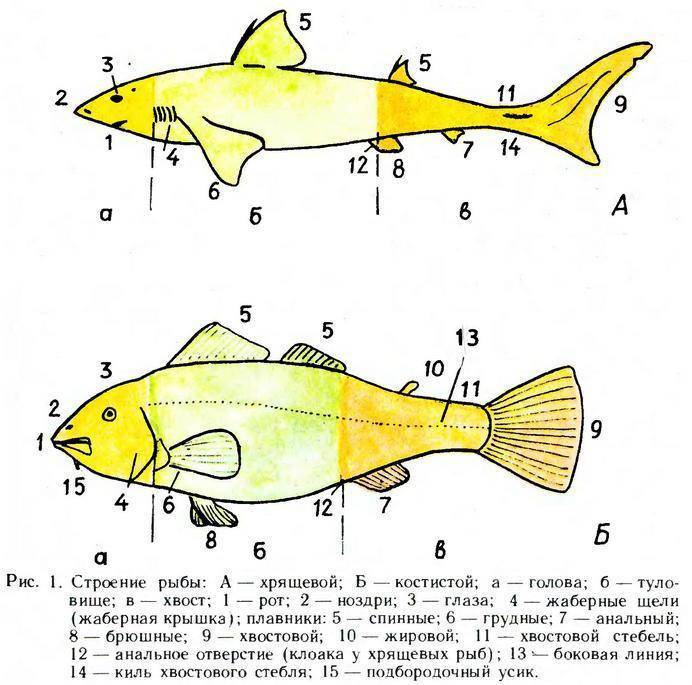
Анатомия рыбы:
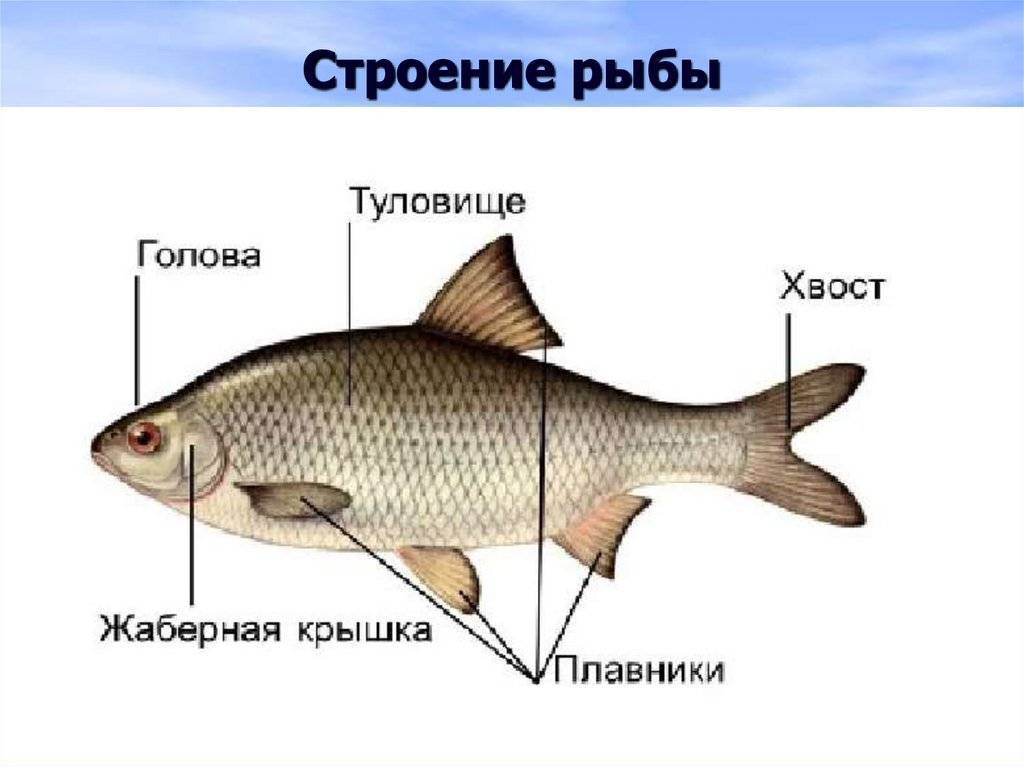
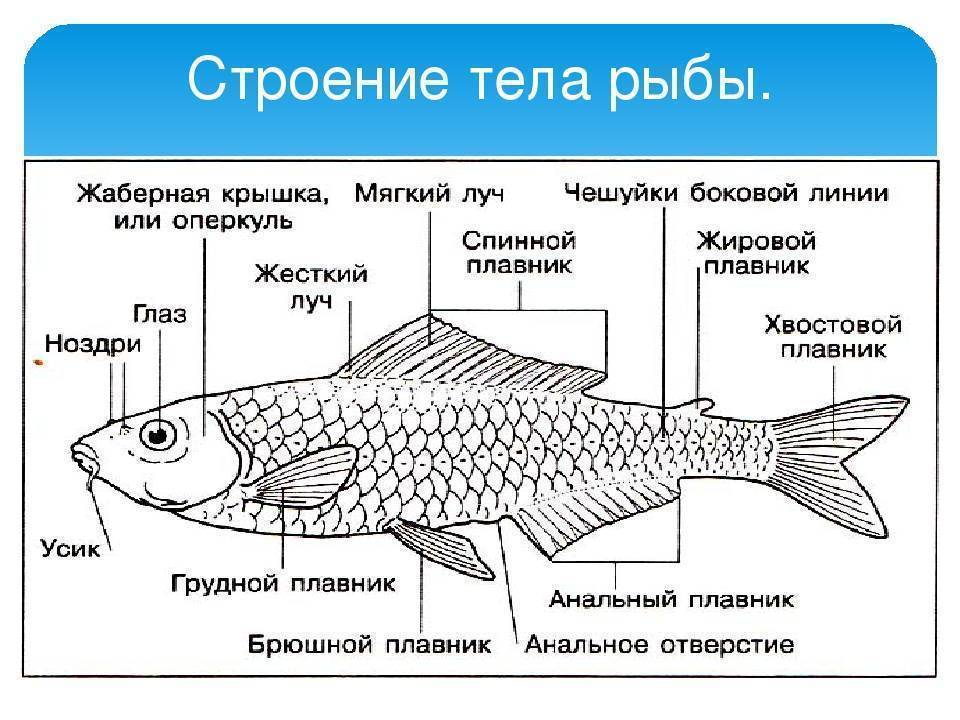
- Тело. Обтекаемое, приплюснуто с боков, что позволяет преодолеть сопротивление водных потоков. Состоит из головы, туловища и хвоста.
- Голова. В верхней части расположены глаза, средней – ноздри, а нижней – рот.
- Покров. Тонкий, пронизан кожными железами, выделяющими слизь. Такая смазка необходима для лучшего скольжения в воде. Защищен чешуйчатым покрытием.
- Жаберные крышки. Прикрывают жаберные щели.
- Плавники. Парные – грудные и брюшные, расположены симметрично по бокам. Непарные – спинные, анальный хвостовой. Локализированы в верхней и задней части туловища.
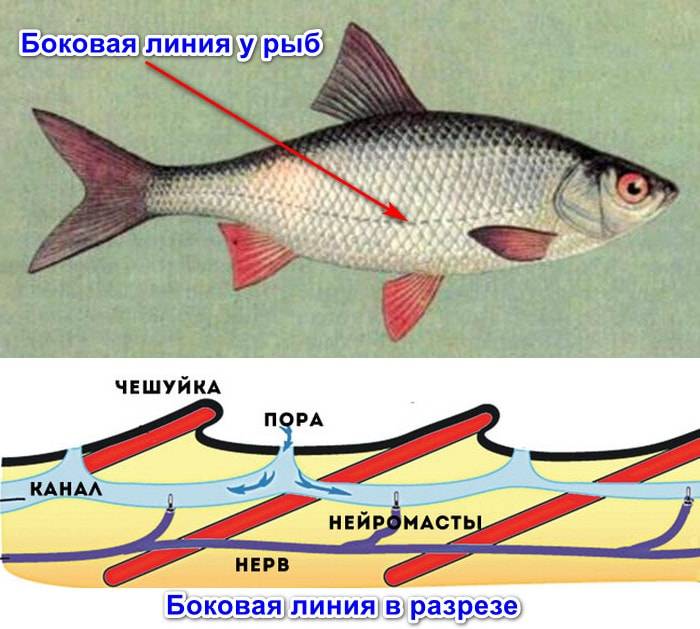
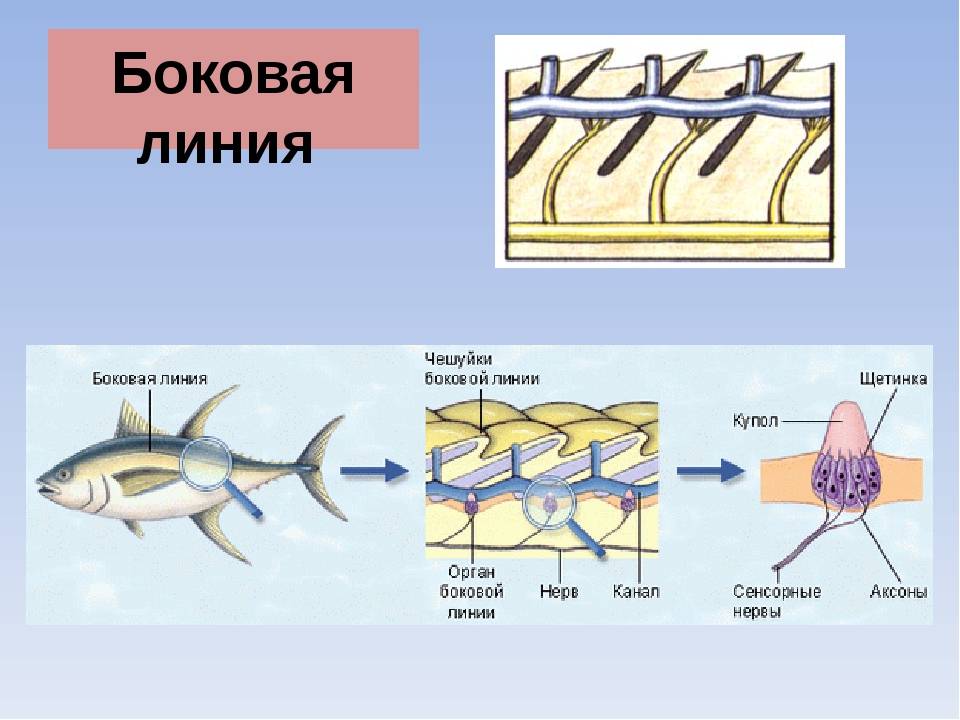
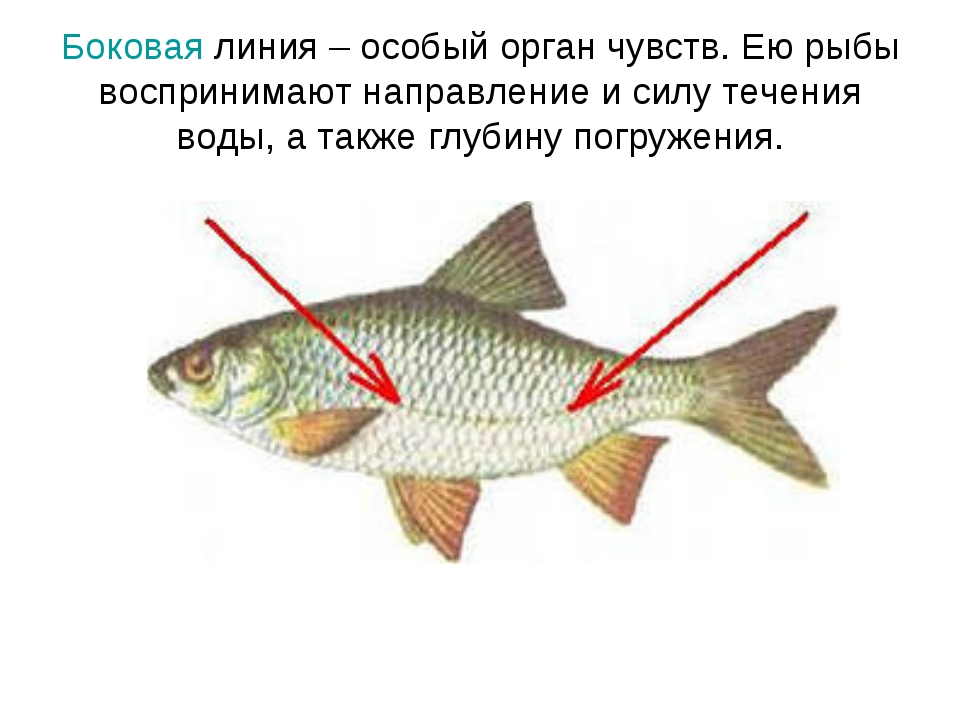
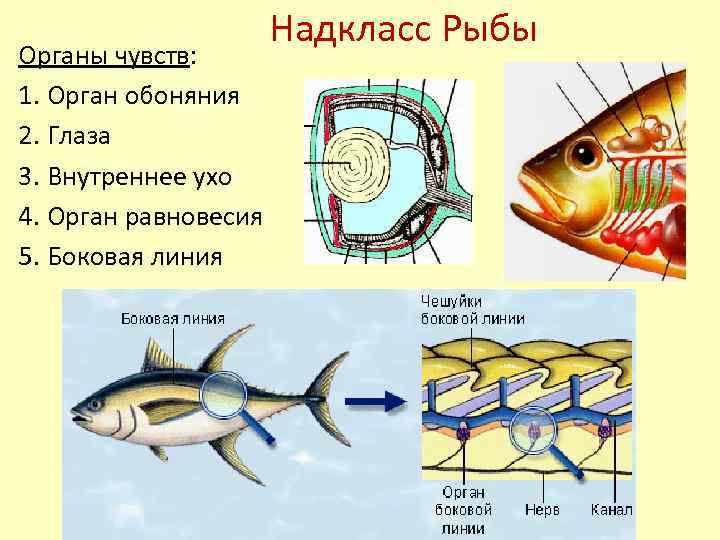
Боковая линия – чешуя и нервные окончания
Боковая линия предназначена для определения направления потоков воды, силы течения. Является значимым органом чувств. Состоит из выходящих на поверхность каналов, на дне которых залегают нервные окончания. Позволяет рыбе ориентироваться в водной среде, ловить добычу, совершать плавательные маневры.
Разновидности чешуи:
- Костная. Встречается у окуневых, сельдевых и карповых, состоит исключительно из костной ткани.
Боковая линия помогает определить направление потоков воды
- Ганоидная. Ромбовидная, частички плотно прилегают друг к другу, образуя панцирь. Содержит ганоин в верхней части и костные структуры внизу. Покрывает тело кистеперых рыб и хвост осетровых.
- Плакоидная. Чешуйки в форме ромба, имеют выступ похожий на шип. В составе дентин и витродентин на выростах. Обладают полостью, которая заполняется соединительным веществом, сосудами или нервными волокнами. Характерна для скатов, акул и хрящевых представителей.




Пищеварительная система.
На переднем конце головы расположен рот. Как правило, верхний рот бывает у рыб, держащихся у поверхности воды; нижний рот характерен для донных рыб; конечный рот — для рыб средних слоев воды. У многих рыб в ротовой полости имеются зубы из дентина, покрытого эмалью. У некоторых видов рыб имеются глоточные зубы. Язык представляет собой мускулистый вырост дна ротовой полости. Ротовая полость переходит в глотку, по бокам которой находятся жаберные щели, далее следует короткий и широкий пищевод, переходящий в желудок.
Объемистый, часто изогнутый в виде колена, желудок переходит в тонкую кишку (карповые рыбы желудка не имеют). У многих рыб на границе желудка и кишки расположены слепые пальцевидные (пилорические) отростки, служащие для увеличения пищеварительной поверхности. Кишка образует петли и открывается наружу анальным отверстием.
Из пищеварительных желез хорошо выражена объемистая печень. Вырабатываемая печенью желчь собирается в желчном пузыре, откуда она поступает в тонкую кишку.
В петле кишки расположена селезенка — кроветворный орган. У большинства рыб между кишкой и позвоночником (на спинной стороне полости тела) расположен плавательный пузырь, наполненный смесью газов и играющий роль гидростатического органа. При увеличении объема плавательного пузыря удельный вес тела рыбы уменьшается рыба поднимается к поверхности; при уменьшении объема — удельный вес увеличивается, рыба опускается.
Пищеварительная система у рыб
Начинается пищеварительная система крупным ртом, который расположен в передней части головы и вооружен челюстями. Имеются крупные мелкие зубы. Позади ротовой полости находится полость глотки, в которой можно увидеть жаберные щели, которые разделены межжаберными перегородками, на которых расположены жабры. Снаружи жабры прикрываются жаберными крышками. Далее находится пищевод, за которым следует достаточно объемистый желудок. За ним расположена кишка.
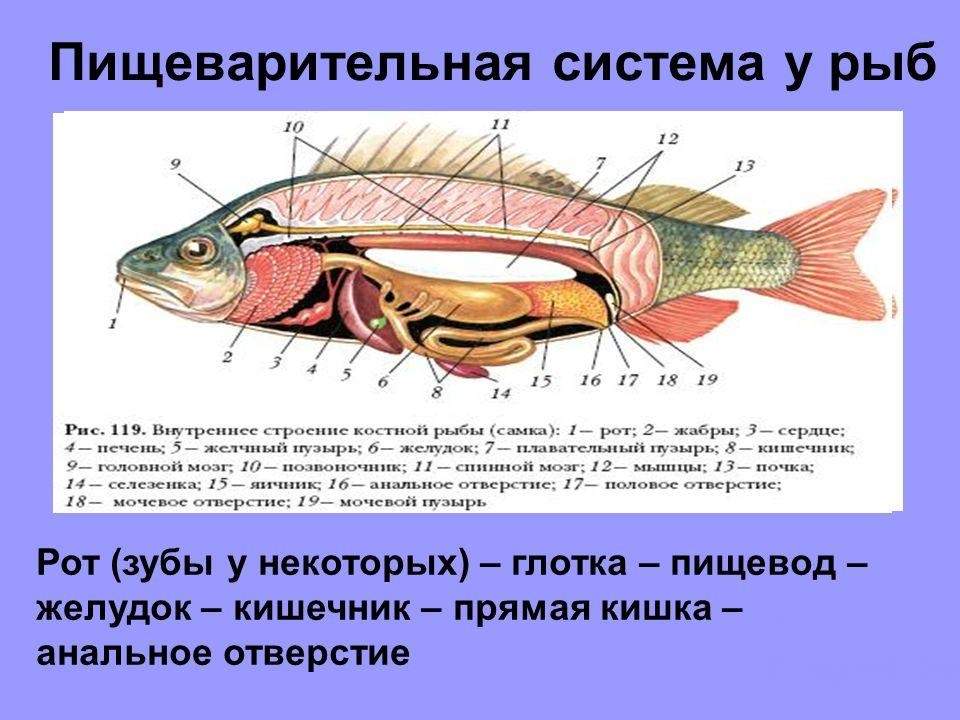 Строение пищеварительной системы рыб.
Строение пищеварительной системы рыб.
Желудок и кишка, используя действие пищеварительных соков, переваривают пищу, причем в желудке действует желудочный сок, а в кишечнике сразу несколько соков, которые выделяют железы стенок кишечника, а также стенки поджелудочной железы. Также участвует в этом процессе и поступающая из печени и желчного пузыря желчь. Переваренная в кишечнике вода и пища всасываются в кровь, а непереваренные остатки через анальное отверстие выбрасываются наружу.
Особым органом, который имеется только у костных рыб, являются плавательный пузырь, который находится под позвоночником в полости тела. Плавательный пузырь возникает в ходе эмбрионального развития в качестве спинного выроста кишечной трубки. Чтобы пузырь был заполнен воздухом, только что появившиеся на свет малек всплывает на поверхность воды и заглатывает воздух в свой пищевод. Спустя некоторое время связь между пищеводом и плавательным пузырем прерывается.
 В природе существуют даже прозрачные рыбы.
В природе существуют даже прозрачные рыбы.
Представляет интерес, то, что некоторые рыбы используют плавательный пузырь в качестве средства, с помощью которого они усиливают издаваемые ими звуки. Правда у некоторых рыб, плавательный пузырь отсутствует. Обычно это те рыбы, которые обитают на дне, а также те, для которых характерны вертикальные быстрые перемещения.
Благодаря плавательному пузырю, рыба не тонет под своей тяжестью. Состоит этот орган из одной или из двух камер и заполняется смесью газов, которая по своему составу близка к воздуху. Объем содержащихся в плавательном пузыре газов может изменяться при поглощении выделении оных через кровеносные сосуды стенок плавательного пузыря, а также при заглатывании воздуха. Таким образом, удельный вес рыбы и объем ее тела и может изменяться в ту или иную сторону. Плавательный пузырь обеспечивает рыбе равновесие между массой ее тела и действующей на нее на определенной глубине выталкивающей силой.
Внутреннее строение рыб
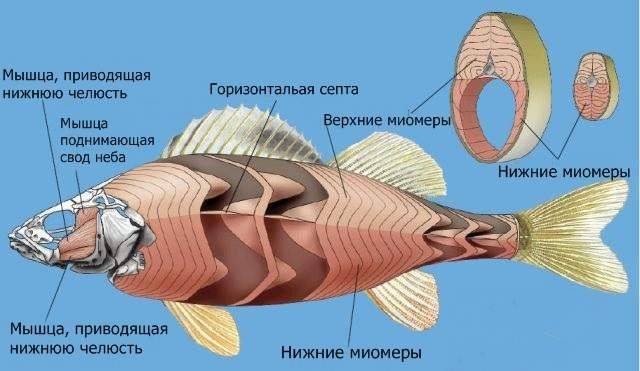
Костно-мышечная система рыб, как и у наземных животных, состоит из мышц и скелета. В основе скелета лежит состоящий из отдельных позвонков позвоночник и череп. У каждого позвонка имеется утолщенная часть, которая называется телом позвонка, а также нижние и верхние дуги. Вместе, верхние дуги образуют канал, в котором и находится спинной мозг, который защищается от травм дугами. В верхнем направлении от дуг отходят длинные остистые отростки. В туловищной части нижние дуги разомкнуты. В хвостовой части позвоночника нижние дуги образуют канал, внутри которого проходят кровеносные сосуды. Ребра примыкают к боковым отросткам позвонков и выполняют целый ряд функций, в первую очередь защиту внутренних органов, и создание необходимой опоры для мускулатуры туловища. Наиболее мощная мускулатура у рыб находится в области хвоста и спины.
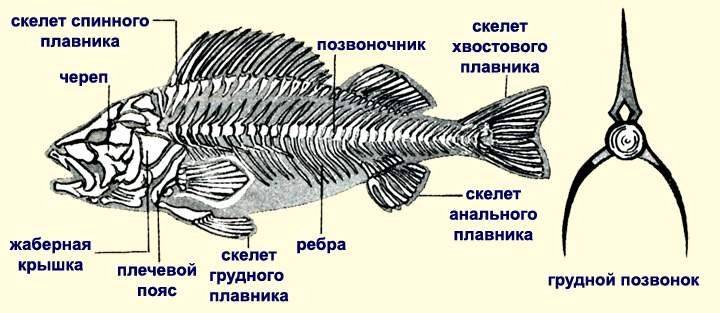 Строение скелета рыбы.
Строение скелета рыбы.
Скелет рыбы включает в себя кости и костные лучи как парных, так и непарных плавников. У непарных плавников скелет состоит из множества крепленных в толще мускулатуры удлиненных косточек. В брюшном поясе находится единая кость. У свободного брюшного плавника скелет состоит из множества длинных косточек.
В скелет головы входит и небольшая черепная коробка. Кости черепа служат защитой для головного мозга, но большую часть скелета головы занимают кости верхних и нижних челюстей, кости жаберного аппарата и глазниц. Говоря о жаберном аппарате, можно отметить в первую очередь жаберные крышки крупного размера. Если жаберные крышки немного приподнять, то под ними можно будет увидеть парные жаберные дуги: левые и правые. На этих дугах размещены жабры.
Что касается мышц, то в головной части их немного они расположены по большей части в районе жаберных крышек, на затылке и челюстях.
 Мышцы рыбы.
Мышцы рыбы.
К скелетным костям прикрепляются обеспечивающие своей работой движение мышцы. Основная часть мышц равномерно расположена в спинной части тела животного. Наиболее развитыми являются мышцы двигающие хвост.



Функции костно-мышечной системы в организме рыб самые различные. Скелет служит защитой для внутренних органов, костные плавниковые лучи защищают рыбу от соперников и хищников, а весь скелет в сочетании с мышцами позволяет этому обитателю вод двигаться и защищаться от столкновений и ударов.





